Bilde 1. Prinsippskisse av øye
| Forfattere (årstall) |
Navn på artikkel |
Publikasjon/utgivelse |
| Aharonowitz Y.og Cohen G.(1981) |
To fold or not to fold |
Science vol.260, ss.1903-1904 |
| Bahill og Stark (1979) |
The trajectories of saccadic eye movements |
Scientific American, vol.240, Jan. |
| Beeman R.W (1982) |
Recent advances in the mode of actionof insectiicides |
Annual Review of Entomology, vol. 27, ss.253-281 |
| Brock R.D. (1980) |
Mutagenesis and Crop Production |
The Biology of Crop Productivity, New York, Ac.Press. ss.383-409 |
| Burleigh B.D. et al, (1974) |
A comparison of wild type an mutant ribitol dehydrogenase fraom Klebsiella aerogenes |
Biochemical Journal vol.143, ss.341-352 |
| Cairns J.J.et al (1988) |
The origins of mutants |
Nature vol.335, ss.142-145 |
| Campell et al. (1973) |
Evolution for a second gene for beta-galactosidase in E-coli |
Proceedings National Academy of Science USA, vol.70, ss1841-1845. |
| Davies et al (1971) |
R-factors Biochemical mechanisms of resistance to aminoglycoside antibiotics |
Annals of the NY Academy of Science, vol. 182 ss.226-233 |
| Davies et al (1972) |
The genetics of bacterial ribosomes |
Annual Review of Genetics, vol.6, ss.203-234 |
| Darnell et al. (1986) |
Molecular Cell Biology |
New York: Scientific Am. Books |
| Gartner, T.K. et E. Orias (1966) |
Effects of mutations to streptomycin resistance on the reate of translation of mutant genetic information |
Journal of Bacteriology, vol. 91, ss.1021-1028 |
| Hall , B.G. (1978) |
Regulation of newly evolved enzymes IV. Directed evolution of the eBetag repressor |
Genetics vol. 90, ss.673-681 |
| Hall , B.G. (1982) |
Evolution in a Petri dish: The evolved Beta-galactosidase system as a model for studying acquisitive evolution in the laboratory, |
Ev. Biology, vol.15.ss.85-150 |
| Hall , B.G. (1988) |
Adaptive mutatitons that requires multiple spontaneous mutations. I. Mutations involving an insertion sequence |
Genetics vol. 120, ss.887-897 |
| Herskowitz I.H. (1962) |
Genetics |
Boston: Little Brown |
| Landau R.L. (1967) |
Endocrinology |
Encyclopedia Brittanica, Vol. 8, ss. 378-382 |
| Kohler, I. (1962) |
Experiments with goggles |
Scientific American, vol.206, Mai |
| Konzak (1977) |
Genetic control of the content, amino acid composition and prosesseing properties of proteins in wheat |
Advances in Genetics, vol.19, ss.407-582 |
| Lerner et al. (1964) |
Evolution of a cathabolic pathway in bacteria |
Science, vol.146, ss.1313-1315 |
| Levine og MacNichol, (1982) |
Color vision in fishes |
Scientific American, vol.246, Febr. |
| Michael C.R. (1969) |
Retina processing of visual images |
Scientific American, vol.220, Mai |
| Pettigrew J.D., (1972) |
The neurophysiology of binocular vision |
Scientific American, vol.227, Aug |
| Rowland M.W, (1987) |
Fitness of insectiicide resistance |
Nature vol. 327, s.194 |
| Schnapf og Baylor (1987) |
How photoreceptor eyes respond to light |
Scientific American, vol.256, April. |
| Stryer, L. (1988) |
Biochemistry (3.utg) |
New York: Freeman |
| Tanaka et al.(1984) |
Picrotoxinin receptor in the central nervous system of American cockroach: its role in the action of cyclodiene-type insecticides |
Pesticide Biochemistry and Physiology, vol. 22, ss.117-127 |
| Wilson, E.O. (1992) |
The Diversity of Life |
Cambridge, Harvard UP |
Stoffutvalg og bilder ved Asbjørn E. Lund
.
 I noen spesielle tilfeller kan en organisme tilpasse seg enkelt ved å droppe foreldet kompleksitet uten å legge til noen ny. Dette ville være et tilfelle av å tape informasjon uten å vinne noen ny. Men om evolusjon fungerte bare på dette viset, kunne den ikke ha bygd opp utvikling av all kompleksitet vi ser omkring oss. Hvordan kunne den ha bygd opp informasjonen den mister? Hvordan kunne f.eks. ha bygd opp et system for syn og øyne (kilder nederst):
I noen spesielle tilfeller kan en organisme tilpasse seg enkelt ved å droppe foreldet kompleksitet uten å legge til noen ny. Dette ville være et tilfelle av å tape informasjon uten å vinne noen ny. Men om evolusjon fungerte bare på dette viset, kunne den ikke ha bygd opp utvikling av all kompleksitet vi ser omkring oss. Hvordan kunne den ha bygd opp informasjonen den mister? Hvordan kunne f.eks. ha bygd opp et system for syn og øyne (kilder nederst):  For at en mutasjon skal spille en rolle i evolusjon, må den innebære en form for fordel for organismen, og den må bli selektert (positiv seleksjonsverdi). Finnes det slike mutasjoner? Ja, det gjør det. Det finnes noen mutasjoner som er kjent for å tjene organismen til fordel i spesielle tilfeller. Vi har allerede sett at en tilbake-mutasjon kan gjenvinne en tapt funksjon.
Det finnes eks. på mutasjoner som gir bakterier resistens mot streptomycin. Det er noen som gir insekter resistens mot DDT. Også har vi alt innen avlsforskning, der anvendt genetikk kan vise at kvantitative trekk kan manipuleres, f.eks. størrelse på tomater, volum av melk hos ei ku, avkastning hos hvete etc. I forskning utført over mange år, har forskere vist at de kan få store økninger i disse trekkene. Men kan noen av disse eks. tjene som prototyp på de små, stegvise mutasjoner som kan bli knyttet sammen i lange sekvenser for å oppnå makroevolusjon?
For at en mutasjon skal spille en rolle i evolusjon, må den innebære en form for fordel for organismen, og den må bli selektert (positiv seleksjonsverdi). Finnes det slike mutasjoner? Ja, det gjør det. Det finnes noen mutasjoner som er kjent for å tjene organismen til fordel i spesielle tilfeller. Vi har allerede sett at en tilbake-mutasjon kan gjenvinne en tapt funksjon.
Det finnes eks. på mutasjoner som gir bakterier resistens mot streptomycin. Det er noen som gir insekter resistens mot DDT. Også har vi alt innen avlsforskning, der anvendt genetikk kan vise at kvantitative trekk kan manipuleres, f.eks. størrelse på tomater, volum av melk hos ei ku, avkastning hos hvete etc. I forskning utført over mange år, har forskere vist at de kan få store økninger i disse trekkene. Men kan noen av disse eks. tjene som prototyp på de små, stegvise mutasjoner som kan bli knyttet sammen i lange sekvenser for å oppnå makroevolusjon? Før vi ser videre på mutasjoner og informasjon, skal vi kikke på begrepet 'spesifisitet'. Dess mer spesifikk et gen er, desto mer informasjon inneholder det. Informasjonen i et gen er det samme som informasjonen i proteinet som det koder for. For å se på et eks. på spesifikk informasjon, kan vi ta for oss ett binært tall. Det har relevans for biologien, som benytter 4 ulike byggeklosser (A,C,T og G), noe som kan uttrykkes ved hjelp av 3 binære siffer: (00,01, 10, 11). I tillegg til de to verdiene (0 og 1), så er plasseringen av sifrene stor betydning. Liksom det desimale, så er det binære tallsystem et posisjonssystem. Vi skal ta et eks. på hvordan en regner om fra binært til desimalt tallsystem: Betrakt det binære tallet 11001. For å finne ut hva det innebærer med 'vanlige' tall, multipliserer vi verdien med vekten av posisjonen, slik figuren illustrerer. Det binære tallet 11001 tilsvarer altså det desimale tallet: 25. Det trenger ingen forklaring at en punktmutasjon, endring av verdien, for ett binært siffer vil gi et ganske annet resultat. Dette har sin parallell i biologisk liv.
Før vi ser videre på mutasjoner og informasjon, skal vi kikke på begrepet 'spesifisitet'. Dess mer spesifikk et gen er, desto mer informasjon inneholder det. Informasjonen i et gen er det samme som informasjonen i proteinet som det koder for. For å se på et eks. på spesifikk informasjon, kan vi ta for oss ett binært tall. Det har relevans for biologien, som benytter 4 ulike byggeklosser (A,C,T og G), noe som kan uttrykkes ved hjelp av 3 binære siffer: (00,01, 10, 11). I tillegg til de to verdiene (0 og 1), så er plasseringen av sifrene stor betydning. Liksom det desimale, så er det binære tallsystem et posisjonssystem. Vi skal ta et eks. på hvordan en regner om fra binært til desimalt tallsystem: Betrakt det binære tallet 11001. For å finne ut hva det innebærer med 'vanlige' tall, multipliserer vi verdien med vekten av posisjonen, slik figuren illustrerer. Det binære tallet 11001 tilsvarer altså det desimale tallet: 25. Det trenger ingen forklaring at en punktmutasjon, endring av verdien, for ett binært siffer vil gi et ganske annet resultat. Dette har sin parallell i biologisk liv. På samme måte kan vi se at ved å redusere spesifisiteten til matchende sekvens i et enzym går hånd i hånd med reduksjon av informasjonen i det. Den lettest kjennelige endring i et enzym, er i dets rolle som katalysator i en reaksjon. Andre type endringer påvirker hvor godt enzymet passer til sitt substrat. En mutasjon nesten hvor som helst i et enzym, som er kontrollert av et allosteric sted (Selve ordet 'allosteric' er et ord som anvendes spesielt om handlemåten til enzymer. Ordet kommer av det greske: allos-'andre' og stereos-'solid'), -vil påvirke noen aspekter av dets ytelse. Laboratorie-forsøk kan ikke påvise alle effektene av en endring på en aminosyre. Bare gjennom nitide granskinger av levende liv hvor enzymet kan uttrykke sin fulle kapasitet, kan en håpe å avlære alle effektene av å endre en aminosyre.
På samme måte kan vi se at ved å redusere spesifisiteten til matchende sekvens i et enzym går hånd i hånd med reduksjon av informasjonen i det. Den lettest kjennelige endring i et enzym, er i dets rolle som katalysator i en reaksjon. Andre type endringer påvirker hvor godt enzymet passer til sitt substrat. En mutasjon nesten hvor som helst i et enzym, som er kontrollert av et allosteric sted (Selve ordet 'allosteric' er et ord som anvendes spesielt om handlemåten til enzymer. Ordet kommer av det greske: allos-'andre' og stereos-'solid'), -vil påvirke noen aspekter av dets ytelse. Laboratorie-forsøk kan ikke påvise alle effektene av en endring på en aminosyre. Bare gjennom nitide granskinger av levende liv hvor enzymet kan uttrykke sin fulle kapasitet, kan en håpe å avlære alle effektene av å endre en aminosyre. Fig.2: Det ferdige Produkt hindrer så vanligvis enzym E1 som katalyserer 1.steg.
Fig.2: Det ferdige Produkt hindrer så vanligvis enzym E1 som katalyserer 1.steg.
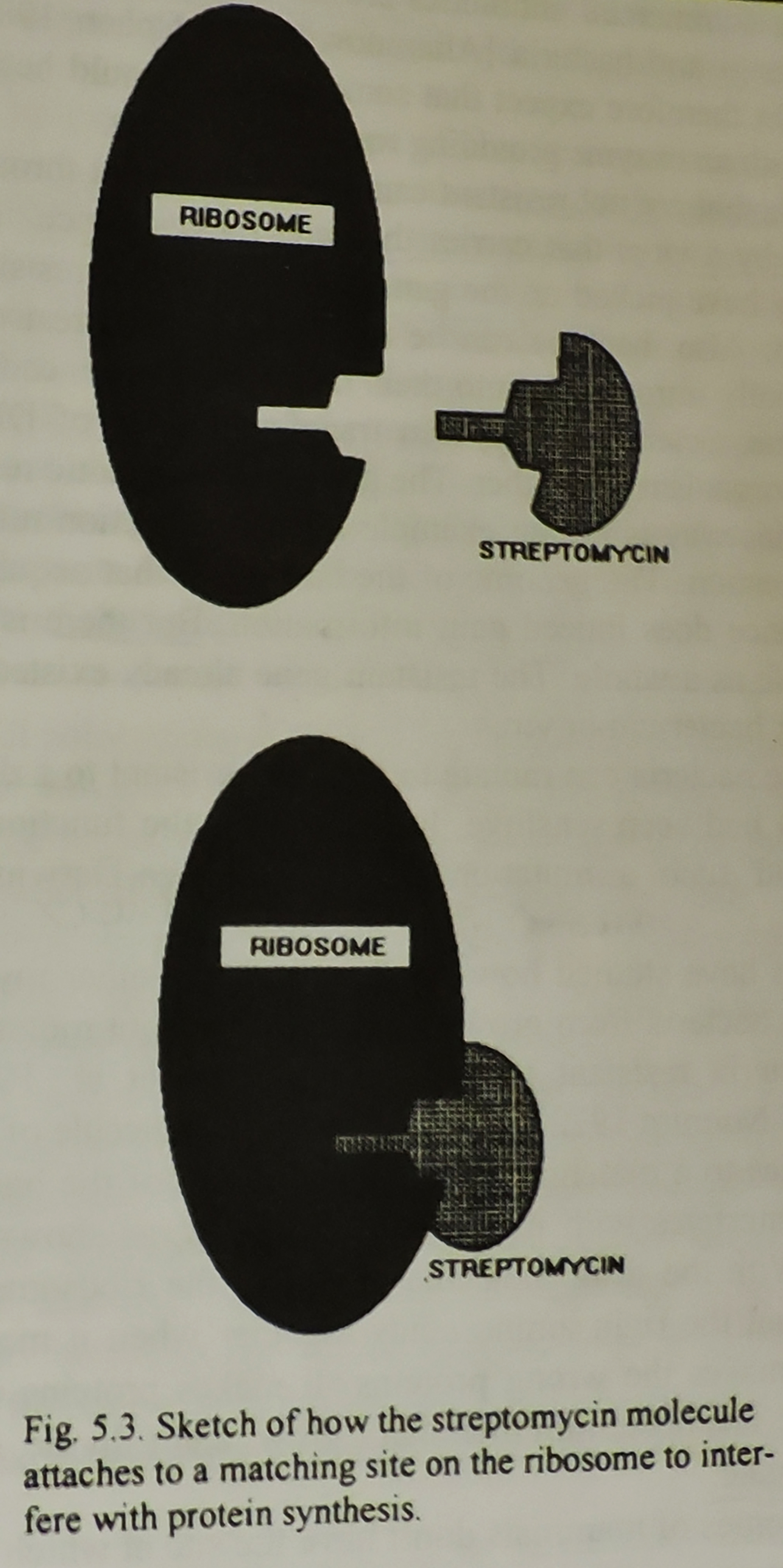 mutasjonen skjer der streptomycinet binder seg til ribosomet, så ville det ikke lenger være noe sted for legemiddelet å knytte seg til (Bilde 5). Legemiddelmolekylet kan ikke lenger knytte seg til ribosomet, så det kan ikke påvirke dets produksjon av protein, og bakterien blir resistent.
mutasjonen skjer der streptomycinet binder seg til ribosomet, så ville det ikke lenger være noe sted for legemiddelet å knytte seg til (Bilde 5). Legemiddelmolekylet kan ikke lenger knytte seg til ribosomet, så det kan ikke påvirke dets produksjon av protein, og bakterien blir resistent.
 substitusjoner av enkelt-nukleotider.
substitusjoner av enkelt-nukleotider. Noen grunner til at den normale cellen ikke kan bruke xylitol har med cellens genetiske kontrollsystem å gjøre. Virkemåten er skissert i Figur 4. Enzymet RDH starter å tilvirkes når genet Y er slått på. Gen Y er normalt AV , men blir slått PÅ i nærvær av ribitol. Molekyler som ribitol kommer ikke lett inn i cellen gjennom celleveggen. De kommer inn om cellen tillater dem adgang. Da benytter cellen en spesiell transport-mekanisme, kjent som et permease enzym, som sitter i celleveggen og slipper molekylet inn. Det foreligger tre problemer som hindrer den normale cellen fra å benytte xylitol. De er:
Noen grunner til at den normale cellen ikke kan bruke xylitol har med cellens genetiske kontrollsystem å gjøre. Virkemåten er skissert i Figur 4. Enzymet RDH starter å tilvirkes når genet Y er slått på. Gen Y er normalt AV , men blir slått PÅ i nærvær av ribitol. Molekyler som ribitol kommer ikke lett inn i cellen gjennom celleveggen. De kommer inn om cellen tillater dem adgang. Da benytter cellen en spesiell transport-mekanisme, kjent som et permease enzym, som sitter i celleveggen og slipper molekylet inn. Det foreligger tre problemer som hindrer den normale cellen fra å benytte xylitol. De er: Selv om X1 ikke bidro noe til å få xylitol inn i cellen, kunne X1 vokse på xylitol, noe den ville typen ikke kunne. Men fordelen ved mutasjonen kom via tap av informasjon, ved å ødelegge undertrykkelses(repressor)-proteinet. X2 familien resulterte i en punktmutasjon i X1 bakterien. Den andre mutasjonen skjer ved posisjon (2) i Fig. 4. Den endret selve enzymet og økte dets aktivitet på xylitol -ca. 2.5 ganger raskere. Det er lett å tenke at siden enzymet ble mer aktivt på xylitol, at det ble mer spesifikt, og at genetisk informasjon økte. Men det viser seg at enzymet ble mer generelt, ved at det virket bedre på et bredere spekter, men mindre spesifikt -dårligere effekt i forhold til ribitol. (Fig. 5). Brian Hartley og hans gruppe ved Imperial College i London gjorde en studie av enzymet (Burleigh et al, 1974). De sammenlignet aktiviteten til X2 mutanten med enzymet i natur-varianten. De målte aktiviteten til enzymet på ribitol, xylitol og L-arabitol, et annet unaturlig substrat. De fant at sammenlignet med den naturlige varianten, var mutant enzymet (X2): a) mindre aktiv på ribitol b) mer aktiv på xylitol og c) mer aktiv på L-arabitol.
Selv om X1 ikke bidro noe til å få xylitol inn i cellen, kunne X1 vokse på xylitol, noe den ville typen ikke kunne. Men fordelen ved mutasjonen kom via tap av informasjon, ved å ødelegge undertrykkelses(repressor)-proteinet. X2 familien resulterte i en punktmutasjon i X1 bakterien. Den andre mutasjonen skjer ved posisjon (2) i Fig. 4. Den endret selve enzymet og økte dets aktivitet på xylitol -ca. 2.5 ganger raskere. Det er lett å tenke at siden enzymet ble mer aktivt på xylitol, at det ble mer spesifikt, og at genetisk informasjon økte. Men det viser seg at enzymet ble mer generelt, ved at det virket bedre på et bredere spekter, men mindre spesifikt -dårligere effekt i forhold til ribitol. (Fig. 5). Brian Hartley og hans gruppe ved Imperial College i London gjorde en studie av enzymet (Burleigh et al, 1974). De sammenlignet aktiviteten til X2 mutanten med enzymet i natur-varianten. De målte aktiviteten til enzymet på ribitol, xylitol og L-arabitol, et annet unaturlig substrat. De fant at sammenlignet med den naturlige varianten, var mutant enzymet (X2): a) mindre aktiv på ribitol b) mer aktiv på xylitol og c) mer aktiv på L-arabitol. X3 ble til fra X2 gjennom en punkt mutasjon ved pkt. (3) i Fig. 6. Denne mutasjonen deaktiverte genet for regulering av det transport-enzymet som frakter et næringsstoff (D-arabitol) til cellen. Transportsystemet viser seg også være i stand til å frakte xylitol inn i cellen, men vanligvis blir ikke transport-enzymet tilvirket om ikke tilhørende gen blir slått PÅ i nærvær av D-arabitol. I fravær av D-arabitol blir genet holdt i tilstand AV ved et repressor protein. Selv om transport-enzymet også kan virke på xylitol, blir det normalt ikke tilvirket, om ikke det blir trigget av D-arabitol. Det som skjedde ved mutasjonen fra X2 til X3, var at mutasjonen deaktiverte genet som kodet for repressor proteinet. Dermed kunne ikke genet til transport-enzymet undertrykkes. Som et resultat ble transport-enzymet laget i store mengder, uten regulering. Dermed var det ikke behov for at det måtte trigges av D-arabitol. Xylitol får dermed en fri reise inn i cellen, ved hjelp av transport-enzymet ment for D-arabitol. Dermed kunne mye mer xylitol entre X3-cellen enn for X1 eller X2. Dermed kunne X3 ernære seg bedre på xylitol enn X2 kunne. Som tidligere for X1 og X2 ble spesifisiteten til et gen redusert, og informasjon gikk tapt.
X3 ble til fra X2 gjennom en punkt mutasjon ved pkt. (3) i Fig. 6. Denne mutasjonen deaktiverte genet for regulering av det transport-enzymet som frakter et næringsstoff (D-arabitol) til cellen. Transportsystemet viser seg også være i stand til å frakte xylitol inn i cellen, men vanligvis blir ikke transport-enzymet tilvirket om ikke tilhørende gen blir slått PÅ i nærvær av D-arabitol. I fravær av D-arabitol blir genet holdt i tilstand AV ved et repressor protein. Selv om transport-enzymet også kan virke på xylitol, blir det normalt ikke tilvirket, om ikke det blir trigget av D-arabitol. Det som skjedde ved mutasjonen fra X2 til X3, var at mutasjonen deaktiverte genet som kodet for repressor proteinet. Dermed kunne ikke genet til transport-enzymet undertrykkes. Som et resultat ble transport-enzymet laget i store mengder, uten regulering. Dermed var det ikke behov for at det måtte trigges av D-arabitol. Xylitol får dermed en fri reise inn i cellen, ved hjelp av transport-enzymet ment for D-arabitol. Dermed kunne mye mer xylitol entre X3-cellen enn for X1 eller X2. Dermed kunne X3 ernære seg bedre på xylitol enn X2 kunne. Som tidligere for X1 og X2 ble spesifisiteten til et gen redusert, og informasjon gikk tapt.